The Gut Terrain: Zonulin, Leaky Barriers & The LPS Cascade
The Medical Matrix has exactly one framework for gut problems: diagnose a condition, prescribe a suppressor. Irritable bowel — antispasmodics. Acid reflux — proton pump inhibitors. Inflammatory bowel — immunosuppressants. Not one of these interventions asks the foundational question: why did the barrier fail? Sovereign bio-physics locates the mechanism precisely — a single gatekeeper protein that determines whether your gut lining is a selective membrane or an open drain. When that gate stays open, it does not just produce digestive symptoms. It generates the chronic low-grade inflammation behind conditions that carry no gut diagnosis at all.

The gut lining replaces itself completely every 4 to 5 days.
This is one of the fastest tissue renewal cycles in the human body. It means the barrier is never static — it is always either being rebuilt correctly or rebuilt incorrectly. Every input that touches the gut lining every day is a vote for one outcome or the other. The terrain model does not ask what is wrong with your gut. It asks what has been attacking the rebuild cycle long enough to make the barrier structurally fail.
The gut barrier collapse operates across four distinct bio-physical stages. Each stage has a different mechanism, a different terrain signature, and a different sovereign intervention point.
Pillar I: The Barrier Architecture — Tight Junctions & The Zonulin Gate
The protein complexes that hold the lining together, the single molecule that opens them, and the triggers that activate it.

The Mechanism
The intestinal epithelium is a single-cell-thick layer lining the gut wall — the thinnest interface between the external environment and the bloodstream. Between each epithelial cell sit protein complexes called tight junctions: assemblies of occludin, claudins, and zonula occludens-1 (ZO-1) that function like molecular zipper locks. When tight, these junctions create a selective barrier — nutrients pass through, pathogens and undigested particles do not.
The gatekeeper of this entire system is a single protein: zonulin. Discovered by gastroenterologist Alessio Fasano in 2000 while researching cholera toxin, zonulin is the only known physiological regulator of intestinal permeability in humans. It works by triggering the disassembly of tight junction protein complexes, temporarily widening the paracellular space between cells. In normal physiology, this is a calibrated, reversible process — zonulin briefly opens to allow immune sampling of gut contents and then closes.
The pathology occurs when zonulin release becomes chronic. Two triggers activate sustained zonulin production beyond all others: gliadin (the protein fraction of gluten, in both celiac and non-celiac individuals) and bacterial dysbiosis — particularly the presence of gram-negative bacteria in the small intestine where they do not belong. Both activate zonulin via the CXCR3 receptor on epithelial cells. When the trigger is removed, tight junctions can close. When the trigger is permanent — through daily gluten consumption in a compromised gut, or through established dysbiosis — the barrier remains structurally open. (Fasano, 2012, PMC3384703)
The Matrix Trap: Gastroenterology tests for gross structural damage — ulcers, polyps, confirmed celiac disease. A chronically open tight junction produces no lesion visible on endoscopy, no antibody flag on standard bloodwork, and no diagnosis. The patient is told their gut is normal. The barrier has been functionally open for years.
Secondary triggers that sustain zonulin production include: NSAIDs (ibuprofen, aspirin — both documented to increase intestinal permeability within hours of ingestion), chronic psychological stress via cortisol-mediated tight junction downregulation, alcohol, and emulsifiers found in processed food (polysorbate-80, carboxymethylcellulose) which disrupt the intestinal mucus layer that sits above the epithelium as the barrier's first line of defence.
The Sovereign Protocol: Seal the Gate
Pillar I Protocol — Restore the Mucosal Layer
Phase 1: Re-establish the Mucus Layer- Slippery Elm bark (Ulmus rubra), 400–500mg powder in water, before meals: The mucilage of slippery elm is a demulcent — it coats and soothes the intestinal mucosa, physically restoring the mucus layer above the epithelium. This creates the protective gel environment in which epithelial repair can proceed. Not a pharmacological intervention — a substrate intervention. Source Slippery Elm →
- Marshmallow Root (Althaea officinalis), decoction, 3–4g dried root in cold water (cold infusion, 8 hours): Cold-infusion preserves the polysaccharide mucilage intact. Marshmallow root's mucopolysaccharides coat the gut lining more deeply than slippery elm, reaching further into the small intestinal crypts where enterocyte regeneration originates. Source Marshmallow Root →
- Eliminate gluten-containing grains for a minimum of 60 days during active barrier restoration — the most consistent zonulin trigger in compromised terrain. This is not a permanent dietary prescription. It is a terrain intervention window.
- Discontinue NSAID use where clinically possible. If anti-inflammatory support is required, botanical alternatives with a clean gut safety profile should be used (see Pillar IV).
Pillar II: Metabolic Endotoxemia — The LPS Cascade
What enters the bloodstream when the barrier opens, how it activates systemic inflammation, and why no standard panel catches it.

The Mechanism
The most consequential passenger through an open gut barrier is not undigested food protein. It is lipopolysaccharide (LPS) — a structural component of the outer membrane of gram-negative bacteria that constitutes roughly 70% of the gut microbiome. LPS is released when gram-negative bacteria die and shed their cell walls. In a healthy, intact barrier, LPS remains in the gut lumen and is processed by the local immune system without entering systemic circulation. In a leaky barrier, LPS passes through the open tight junctions into the portal vein.
LPS binds to TLR4 receptors (Toll-like receptor 4) on immune cells throughout the body — macrophages, monocytes, endothelial cells, adipocytes, and neurons. TLR4 binding activates NF-κB, the master inflammatory transcription factor. The result is a systemic low-grade inflammatory state that Patrice Cani and colleagues at the University of Louvain named metabolic endotoxemia in 2007. Their landmark study showed that plasma LPS concentrations 2 to 3 times above baseline — well below septic levels — were sufficient to drive the full inflammatory phenotype of obesity and insulin resistance in mice, and that the same LPS elevation was documented in humans on a high-fat Western diet. (Cani et al., 2007, PMID 17456850)
The clinical consequences of sustained, low-grade LPS translocation span the full spectrum of modern chronic disease: systemic low-grade inflammation, insulin receptor desensitisation, hepatic fat accumulation, neuroinflammation (LPS crosses the blood-brain barrier in a compromised state), and cardiovascular endothelial damage. These arrive without a gut diagnosis. The gastroenterologist finds no lesion. The rheumatologist sees elevated CRP. The neurologist notes cognitive decline. The cardiologist measures arterial stiffness. All are reading different symptoms of the same upstream event — an open barrier flooding the system with bacterial endotoxin.
The critical insight: LPS-driven inflammation does not produce the acute fever and septic response that clinical training associates with bacterial exposure. It produces a chronic sub-threshold activation — too low to trigger an emergency response, too sustained to allow the terrain to recover. Standard inflammatory markers (CRP, ESR) fluctuate at the margin. The gut barrier is never tested. The mechanism runs undetected for years.
The connection to Lesson 017 (The Histamine Illusion) is direct: the diamine oxidase (DAO) enzyme responsible for breaking down histamine is produced by the same enterocytes that form the gut barrier. When the epithelium is damaged and barrier integrity is lost, DAO output falls in parallel. Histamine accumulates not because more is being produced — but because the same tissue damage that opened the barrier has eliminated the primary clearance mechanism. Histamine overflow and leaky gut are two expressions of a single epithelial failure.
The Sovereign Protocol: Reduce the LPS Burden
Pillar II Protocol — Intercept the Endotoxin Load
Bind and Clear Circulating LPS- Chlorella (broken cell wall, 3–5g daily): Chlorella's cell wall polysaccharides have documented LPS-binding activity — adsorbing endotoxin in the gut before it can translocate, and supporting its clearance from circulation. Simultaneously supports heavy metal chelation, which accumulates in dysbiotic gut terrain and further disrupts the epithelial barrier. Source Chlorella →
- Turmeric (Curcuma longa, 500–1000mg standardised to 95% curcuminoids, with black pepper or fat): Curcumin is one of the most studied botanical NF-κB inhibitors. It directly blocks the TLR4/NF-κB signalling pathway that LPS activates — reducing the downstream inflammatory cascade at the receptor level. Without fat or piperine, curcumin bioavailability is negligible. Source Turmeric →
- Milk Thistle (Silybum marianum, 140mg silymarin 3× daily): The liver is the first organ to receive portal blood from the gut and bears the initial LPS load. Silymarin protects hepatocytes from LPS-induced damage, supports bile production (which traps LPS for faecal excretion), and suppresses hepatic NF-κB activation. The liver cannot clear what the gut keeps sending — but it can be protected while the upstream barrier is being repaired. Source Milk Thistle →
Pillar III: The Microbiome as Terrain Signal
How the microbial ecosystem governs barrier integrity through butyrate production — and what dysbiosis does to the electrical fuel of the colon.
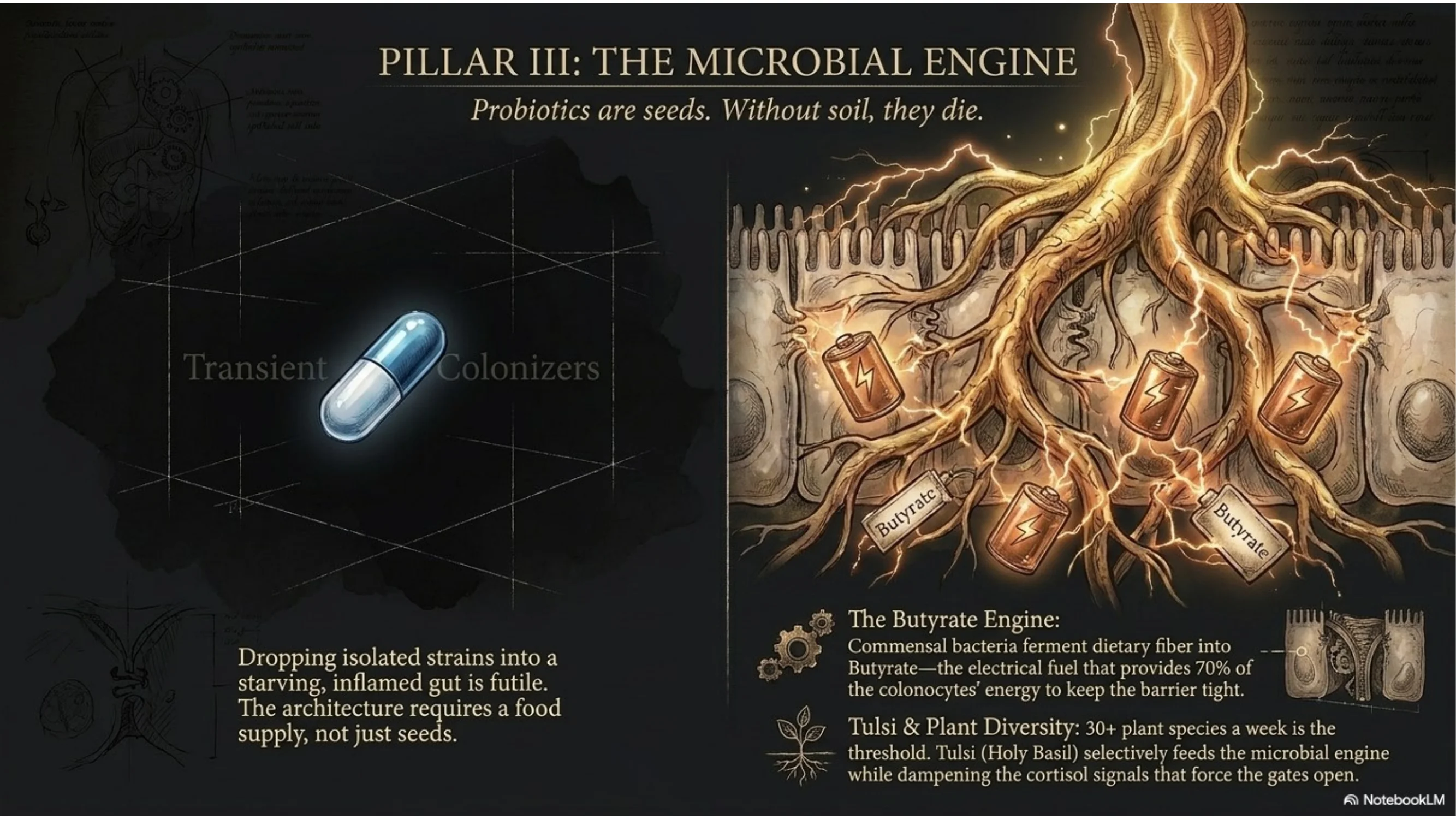
The Mechanism
The gut microbiome is not a passenger in the gut terrain — it is a primary architect of barrier integrity. The connection runs through short-chain fatty acids (SCFAs): metabolic byproducts produced when commensal bacteria ferment dietary fibre in the colon. The three dominant SCFAs — butyrate, propionate, and acetate — have distinct roles in terrain maintenance, but butyrate is the one on which the barrier depends most directly.
Butyrate is the primary energy source of colonocytes — the epithelial cells of the colon. It provides approximately 70% of the total energy requirement of the colonic epithelium. Without adequate butyrate, colonocytes enter a semi-starved state, turnover slows, and barrier integrity degrades from the energetic deficit alone — independently of any inflammatory insult. Butyrate also upregulates the expression of tight junction proteins (claudin-1, occludin) directly, and suppresses NF-κB in colonocytes, reducing local inflammatory signalling that would otherwise degrade the barrier further. (Peng et al., 2009, PMC2683481)
Dysbiosis — the disruption of the microbial ecosystem — preferentially decimates butyrate-producing bacteria: Faecalibacterium prausnitzii, Roseburia intestinalis, and Eubacterium rectale are among the most butyrate-productive species in the human colon and are consistently reduced in inflammatory bowel disease, obesity, type 2 diabetes, and depression. They are sensitive to antibiotics, emulsifiers, and low-fibre diets. As they decline, gram-negative LPS-producing bacteria fill the niche. The microbiome shifts from a butyrate-generating, barrier-protecting ecosystem to an endotoxin-generating, barrier-destroying one.
The pharmaceutical response to dysbiosis is typically a probiotic supplement — isolated bacterial strains delivered in a capsule. The terrain model recognises the limitation: probiotic strains are transient colonisers. Without a substrate — dietary fibre that feeds the resident microbiome — they pass through without establishing. The architecture requires a food supply, not just seeds.
The Sovereign insight: You cannot restore the microbiome without restoring the diet that feeds it. The diversity and butyrate capacity of the gut microbiome directly correlates with dietary plant diversity — not probiotic brand diversity. Thirty different plant species per week is the threshold above which microbial diversity measurably increases. Below that threshold, no supplement combination closes the gap. The terrain responds to substrate, not to isolated strains.
The Sovereign Protocol: Rebuild the Microbial Foundation
Pillar III Protocol — Restore the Butyrate Architecture
Feed the Butyrate Producers- Increase dietary plant diversity to 25–30 species per week as the primary intervention. Legumes, root vegetables, leafy greens, seeds, and herbs all contribute distinct fermentable fibres that feed different microbial niches. This is not a supplement — it is the foundational substrate without which all other interventions are partial.
- Tulsi / Holy Basil (Ocimum tenuiflorum, 300–600mg or 2–3g dried leaf as tea): Documented prebiotic activity — tulsi's polyphenols selectively feed Lactobacillus and Bifidobacterium species while suppressing pathogenic overgrowth. Its adaptogenic action simultaneously reduces the cortisol-driven tight junction downregulation that sustained stress produces. It addresses both the microbial substrate and the stress-axis driver of barrier failure simultaneously. Source Tulsi →
- Eliminate emulsifiers (polysorbate-80, carboxymethylcellulose) from the diet — both are documented to thin the intestinal mucus layer and shift the microbiome toward LPS-producing species within two weeks of consumption.
- Reserve antibiotics for genuine bacterial infections with appropriate culture confirmation. Each antibiotic course produces microbiome disruption that takes 6–24 months to partially resolve, and that never fully returns to the pre-antibiotic baseline in some individuals. The terrain cost of unnecessary antibiotic use is not theoretical — it is measurable in butyrate capacity for years afterward.
Pillar IV: The Full Restoration Stack — Closing the Gate & Rebuilding the Junction
The sequence, the timing, the botanical compounds that directly tighten tight junction proteins.

The Mechanism
Barrier restoration requires more than removing the triggers and feeding the microbiome. The tight junction proteins themselves — occludin, claudins, ZO-1 — are structural molecules whose expression and assembly can be directly upregulated by specific botanical compounds. This is the level at which sovereign restoration operates: not suppressing inflammation downstream, but rebuilding the molecular architecture of the barrier itself.
The compound with the strongest documented direct effect on tight junction protein expression is quercetin — a flavonoid found in onions, capers, and many medicinal herbs. Quercetin inhibits myosin light chain kinase (MLCK), the enzyme that drives the contractile pulling apart of tight junction complexes. By blocking MLCK, quercetin prevents the mechanical disassembly of the junction. It simultaneously upregulates claudin-1 and occludin gene expression, stimulating the synthesis of the proteins the junction needs to rebuild. (Suzuki & Hara, 2011, PMID 21680017)
The restoration sequence matters. Mucosal coating must precede junction tightening — attempting to reassemble tight junctions in an uncoated, inflamed epithelium is like repairing a roof in a storm. The four-phase protocol runs in sequence: coat, reduce inflammation, tighten the junction, rebuild the microbiome substrate. Each phase overlaps with the next, but the order of initiation is non-negotiable.
The Sovereign Protocol: The Gut Terrain Restoration Stack
Pillar IV Protocol — The Gut Terrain Blueprint
Phase 1 (Days 1–14): Coat & Soothe- Slippery Elm 400–500mg in water, 20 minutes before each meal — restore the mucus substrate above the epithelium
- Marshmallow Root cold infusion (3–4g, 8 hours, cold water) — sip throughout the day between meals. Do not heat — polysaccharide mucilage degrades above 40°C
- Remove zonulin triggers: gluten, NSAIDs, emulsifiers
- Turmeric 500–1000mg standardised (with fat + piperine) — NF-κB suppression, LPS cascade interruption
- Chlorella 3–5g daily — LPS binding, heavy metal clearance
- Milk Thistle 140mg silymarin 3× daily — hepatic protection during LPS clearance phase
- Quercetin (500mg, 2× daily with food): Direct MLCK inhibition and claudin/occludin upregulation. Taken with fat for absorption. Minimum 30-day protocol to allow tight junction protein synthesis to accumulate. Source Quercetin →
- Calendula (Calendula officinalis, 300–400mg extract or strong infusion 3×/day): Anti-inflammatory specifically targeted at intestinal epithelial inflammation via triterpenoid (faradiol) activity. Supports epithelial cell regeneration during the tight junction rebuild window. Complements quercetin — different mechanism, same target. Source Calendula →
- Tulsi 300–600mg or 2–3 cups daily as tea — prebiotic action, cortisol modulation, microbiome rebalancing
- Dietary plant diversity: target 25–30 species per week as the long-term butyrate production foundation
- At this stage, if the Phase 1–3 protocol has held, DAO production recovers alongside the epithelium, and the histamine overflow documented in Lesson 017 resolves from its source — not from antihistamine suppression, but from restored barrier architecture
Conclusion: The Gate Was Always There
The gut barrier is 400 square metres of selective intelligence stretched across a single cell layer. It decides, at every moment, what enters the body and what does not. When the architecture of that decision-making fails — when zonulin holds the gate open, LPS floods the portal vein, and butyrate-producing bacteria decline — the downstream consequences land everywhere except on the gut diagnosis that should have been made years earlier.
The Medical Matrix sees obesity, insulin resistance, neuroinflammation, histamine intolerance, joint inflammation, and mood disorders as separate conditions requiring separate medications. Sovereign bio-physics traces all of them to the same upstream address: a barrier system that lost its structural integrity, flooding every downstream terrain with a signal that says — at the cellular level — threat is present, defend, do not restore.
The Sovereign summary:
The gut barrier does not leak because something went wrong inside it. It leaks because the inputs that maintain it — diverse plant fibre, absence of emulsifiers and NSAIDs, a cortisol curve that falls at night — were progressively removed from the modern environment. The terrain lost its maintenance substrate. Restoration is not about treating the gut. It is about restoring the conditions under which the gut can rebuild itself — in the sequence the biology requires, not all at once, and not with suppression.
Close the gate. Clear the load. Feed the builders. The architecture does the rest.